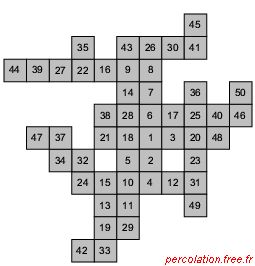
Dans la situation (a) la date d'incorporation est fournie par le nombre précisé sur le site. Dans la situation (b) l'information est fonction du niveau de gris : plus le site est de couleur gris clair, plus sa présence dans l'amas est récente.
Selon cet algorithme, à chaque période, un site inactif voisin d'un site de l'amas, est choisi de façon aléatoire et devient à son tour élément de l'agrégat. À chaque période t, les pondérations affectées aux configurations possibles sont définies de façon à ce que leur somme soit égale à l'unité. On pose comme point origine, le site actif au coin sud-ouest du réseau. Le poids statistique de la configuration à t = 0 est par conséquent unitaire. À t = 1, deux configurations sont possibles, ce qui entraîne une pondération de 1/2 pour chacune. Enfin, aux périodes t = 2 et 3, la pondération passe à 1/4 car quatre possibilités sont concurrentes.
Sous l'hypothèse que le développement s'effectue par le plus proche voisinage, on peut alors conclure sur l'éventualité de certaines chroniques. Sur la figure 2.18 (b), les dates d'incorporation de chaque élément sont compatibles avec l'hypothèse de croissance alors que sur la figure 2.18 (c), elles sont contradictoires. Le site intégré à la date 2 doit par exemple être un plus proche du site origine 1, ce qui n'est pas le cas sur la figure (c).
Sur la figure 2.19 (b) l'effet est plus radical, car dans cette situation l'amas va s'enfermer par son propre développement, conduisant inévitablement à la fin de la croissance. Ce phénomène est qualifié de « piégeage » (trapping).
Dans cet exemple, la croissance de l'amas s'achève à la dixième couronne, car il n'existe plus de sites actifs reliés à ceux de la neuvième. Dans ce cas, la progression n'a pas été limitée par la taille du réseau. L'arrêt est survenu parce qu'aucun site ne permettait de continuer. De la sorte, lorsque les amas ne touchent pas les bords du réseau, les effets de taille finie sont éliminés (Herrmann, 1986, p. 179)26.
À la période suivante, un site est sélectionné au hasard parmi ces quatre candidats (figure 2.21 (b)). Le périmètre se modifie en conséquence et sa taille varie. Ici, le nombre d'éléments parmi lesquels il va falloir choisir un nouveau site augmente. De cette façon, l'amas a une trajectoire qui évolue au gré de l'aléa avec lequel les nouveaux sites sont sélectionnés (figure 2.21 (c) et (d)).
À ce stade, il peut exister de fortes différences entre les résultats géométriques de deux simulations. Au bout de 2 000 itérations, l'amas est relativement plus compact mais il possède toujours un aspect fortement accidenté à sa périphérie (figure (b)). Sur la figure (c), l'amas montre une forme plus homogène. Quelques excroissances sont toujours visibles, mais elles sont modestes relativement à la taille de l'amas.
Poursuivant ce raisonnement, lorsque le nombre d'itérations tend vers l'infini, tous les sites à l'intérieur deviennent actifs. Par conséquent, l'amas est « compact », c'est-à-dire que sa densité intérieure est de 100 % (Herrmann, 1986, p. 158).
Le modèle d'Eden montre ainsi les régularités géométriques de son développement. De la sorte, lorsque l'on compare cet amas avec le résultat particulier d'une simulation (figure 2.22 (c)), la frontière manifeste un aspect beaucoup moins fondamental que l'intérieur de l'amas.
À la période suivante, un site du périmètre est sélectionné au hasard. Pour déterminer si ce site va appartenir à l'amas, un nombre est tiré au sort entre 0 et 1. Sa valeur est comparée à la valeur de p fixée au départ. Si elle est inférieure, le site sélectionné devient actif, sinon le site est neutralisé : «
 »27. L'opération est ensuite réitérée. Sur la
figure 2.25 (b) par exemple, le site candidat est neutralisé alors que
sur la figure 2.25 (c), il est activé. Au fur et à mesure du
développement, le périmètre évolue : les sites neutralisés sont supprimés du périmètre et
les plus proches voisins des nouveaux sites actifs sont ajoutés. L'effectif du périmètre
peut ainsi augmenter ou diminuer d'une période à l'autre
(figure 2.25 (d)).
»27. L'opération est ensuite réitérée. Sur la
figure 2.25 (b) par exemple, le site candidat est neutralisé alors que
sur la figure 2.25 (c), il est activé. Au fur et à mesure du
développement, le périmètre évolue : les sites neutralisés sont supprimés du périmètre et
les plus proches voisins des nouveaux sites actifs sont ajoutés. L'effectif du périmètre
peut ainsi augmenter ou diminuer d'une période à l'autre
(figure 2.25 (d)).Deux représentations sont proposées. La première spécifie les diverses catégories de sites (figure (a)). Les sites actifs sont en gris, les sites neutralisés sont représentés par des «
»,
et ceux du périmètre sont symbolisés par des « · ». Ces derniers étant
susceptibles de prolonger la croissance, il sont également appelés « sites de croissance
» (growth sites). La seconde représentation ne reprend que les sites actifs,
c'est-à-dire l'amas tel qu'il apparaît à la fin (figure (b)).Sur la figure (a), la virulence de la maladie est faible : p = 0,45. Au bout de 87 itérations, l'amas se trouve entouré par une bordure de sites neutralisés. La croissance s'arrête d'elle-même. La taille de l'amas est de 40 sites en plus de l'origine. De façon identique, dans le cas où p = 0,5 et p = 0,55 l'évolution s'annule, respectivement, après 198 et 1 573 itérations. La taille des amas obtenus est de 97 sites en plus de l'origine dans le premier problème (figure (b)) et 871 sites dans le second (figure (c)).
Mise à part le cas où la taille maximale du réseau est fixée, la seule limite au développement de l'amas correspond au nombre d'itérations effectuées pendant la simulation. Si l'algorithme était répété une infinité de fois, la taille de l'amas tendrait alors vers l'infini.
Au fur et à mesure que la valeur de p augmente, l'amas final a une croissance de plus en plus centrée par rapport au site d'origine, car il rencontre de moins en moins de résistance. Au seuil, l'amas est très lacunaire. Des trous de différentes tailles existent sur l'ensemble de la structure (figure (a)). Lorsque p s'élève, la densité de l'amas augmente. À p = 0,7 la structure est nettement moins lacunaire, mais il subsiste tout de même de nombreux trous de petites tailles (figure (b)). Lorsque p atteint 90 %, la structure de l'amas est très dense. Le nombre de trous est faible et leur taille réduite (figure (c)). Pour p = 1, le problème correspond au modèle d'Eden et la structure devient compacte30.
Alors qu'au bout de 500 itérations, l'amas montre une certaine concentration aux environs de l'origine (figure (a)), un étirement relativement important apparaît 1 500 itérations plus tard (figure (b)). Ce développement totalement imprévisible se confirme sur la figure (c) où de nouvelles excroissances se sont ajoutées. Ainsi, dans le quart nord-est, des sites qui dès le départ auraient pu porter la croissance ne sont utilisés qu'après avoir épuisé tous les sites de résistance inférieure. Ce type de croissance amène par conséquent a des résultats de forme variées et imprévisibles. La figure 2.31 illustre la diversité géométrique des amas obtenus avec le modèle d'invasion, après un nombre important d'itérations.
Par le biais de simulation de Monte-Carlo, D. WILKINSON et J.F. WILLEMSEN ont étudié la structure des amas du modèle d'invasion. Dans le cas avec absence de piégeage, la correspondance avec le modèle standard de percolation est clairement mise en évidence. Intuitivement, le problème consiste à activer tous les sites d'un réseau dont le nombre aléatoire associé est inférieur ou égal à la valeur de p*, tel que p* soit la valeur minimale nécessaire pour que l'amas percole. Le modèle d'invasion correspond alors à un sous ensemble des points qui sont actifs dans le modèle de percolation (Wilkinson et Willemsen, 1983, p. 3375). Les valeurs du seuil estimées dans le cas de l'invasion sont alors très proches des valeurs du seuil de percolation (tableau 2.1). Les valeurs du seuil de percolation sont précisées à titre de comparaison33.
Cependant, même si les résultats géométriques obtenus par le modèle d'invasion sont proches de ceux d'un problème de percolation, il existe tout de même des différences (Stauffer et Aharony, 1992, p. 135). Dans le modèle d'invasion, l'amas percole toujours à travers le réseau et par conséquent, il n'existe pas d'amas de taille finie. De plus, le problème ordinaire de percolation se réalise dans un contexte statique alors que le modèle d'invasion est un processus dynamique.